Auditory evoked potentials can be recorded from all regions of the ascending auditory pathway, from cochlear microphonics and cochlear nerve action potentials at the periphery to central auditory cortical responses.
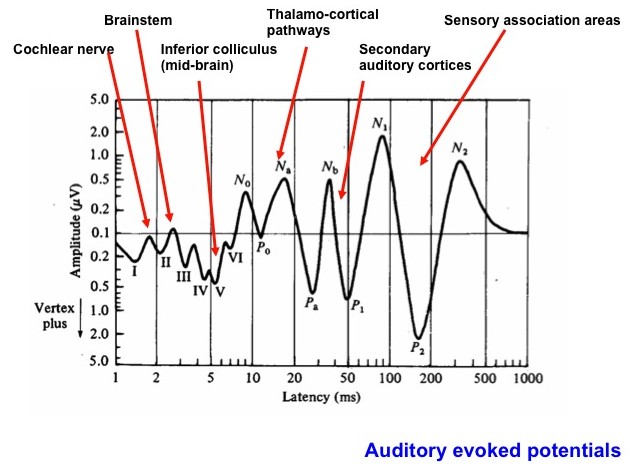
The very pleasant depiction by Netter, shown upper right, provides a simplified overview of the ascending auditory pathway (from one ear). In the lower left panel (based on a figure from Dr. Terry Picton) illustrates the various types of auditory evoked potential that can be obtained over time starting with cochlear events, then auditory brainstem and midbrain to thalamic and cortical potentials. Note the logarithmic time axis. Typically, an evoked potential is generated by a brief click or tone pip from an acoustic transducer such as an insert earphone or headphone. The elicited voltage waveform response is measured through skin electrodes.
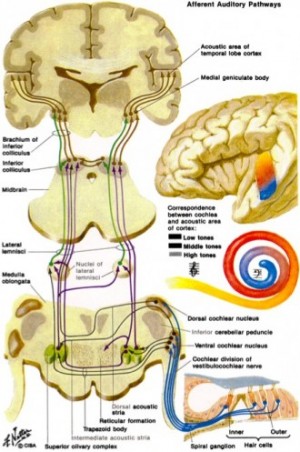
We have measured this whole range of evoked potentials in our lab, but most useful for many research and clinical purposes is the auditory brainstem evoked response (ABR). Below we show a recent study in which we have used ABR recordings in the mouse to study the time course of excito-toxicity after cochlear damage.
As a routine, in animal models we use ABR recording to objectively determine the audiogram of a subject. In this case the ABR audiogram is the threhold response of ABRs generated by tone pip stimuli across frequency.
We have also used ABR recording extensively in clinical studies in children with cochlear implants. In this case the stimulus evoking the auditory response is an electrical pulse through the implanted cochlear electrode (for references see our section on “Translational Research”).
Time-course of cochlear injury discharge (excito-toxicity) determined using ABR measurement.
The dynamics of cochlear excitotoxicity can be monitored from effects on the contralateral ear. After unilateral mechanical ablation of the cochlea (in a mouse model) we observed immediate elevations in auditory brainstem evoked response (ABR) thresholds in the contralateral ear. Threshold elevations peaked at 2 to 3 hours post ablation, and returned to baseline levels after 5 to 6 hours. These contralateral effects are initiated by cochlear afferent injury discharges most likely activating the olivocochlear efferent system.
Graph A shows the rapid increase in thresholds after cochlear ablation at 4kHz frequency, with the difference in thresholds between the experimental and control mice in the initial 5 hours after damage was significant.
Graph B again shows the increase in hearing thresholds after cochlear ablation at 32kHz frequency. This indicates that any injury discharge has a global effect on the cochlear, affecting a range of frequencies along the length of the cochlear.
Six hours after cochlear injury, ABR thresholds were fully returned to pre-lesion baseline levels and remained normal for up to 10 days of monitoring. We have confirmed that our cochlear ablation procedure increases short-term activity levels in the auditory brainstem and midbrain using c-fos labelling.
The study provides insight into the dynamics of glutamate excitotoxicity, a pathological process directly related to acute tinnitus after acoustic trauma, and more generally implicated in many types of brain injury and neuro-degenerative disease.


Negandhi, Harrison, Allemang and Harrison (2014) Time course of cochlear injury discharge (excitotoxicity) determined by ABR monitoring of contralateral cochlear events.
References
Negandhi J, Harrison A.L., Allemang C and Harrison R.V. Time course of cochlear injury discharge (excitotoxicity) determined by ABR monitoring of contralateral cochlear events. Hearing Research. (2014) 315: 34-39.
Grande G., Negandhi J., Harrison R.V. and Wang L.Y. Remodelling at the calyx of Held-MNTB synapse in mice developing with unilateral conductive hearing loss. J Physiol 592.7 (2014) pp 1581-1600.
Gordon K.A., Papsin B.C. and Harrison R.V. Auditory brainstem activity and development evoked by apical verses basal cochlear implant electrode stimulation in children. Clinical Neurophysiology. (2007) 118: 1671-1684.